PRPGの必要性
世界中の作物の損失は、主に土壌伝染病によって引き起こされています。www.modernagriculturefarm.com世界の作物生産性は、植物病原菌によって引き起こされる病気によって悪影響を受けます。 その結果、さまざまな穀物やマメ科作物で毎年20〜40パーセントの収量損失が発生します。
2016-17年の農業シーズンの間に、 57、 インドでは、植物病原菌や害虫から保護するために、000メートルトンの合成農薬が使用されました。しかし、 生物農薬はわずか6340メートルトンしか占めていません。
に似ている 雑草除草剤耐性 、 現代の農業における農薬の継続的な使用と野菜中の残留農薬の存在により、多くの農薬耐性菌が出現しました。 穀類、 穀物も人間の健康に大きな危険をもたらします。
植物の病気と戦うために利用される農薬は、有益な自然の昆虫に影響を与えます、 土壌の肥沃度と土壌の微生物相に悪影響を与えるKhatoonet al。、 2020
さらに、 化学農薬と 肥料 規制されていない無差別な方法で使用されると、土壌の汚染を引き起こし、 水、 と空気、 土壌動物相と微生物相の減少と同様に。世界中の土壌伝染病を防除するために、 合成農薬には多額の費用がかかります。合成化学物質の環境への悪影響により、 植物病害防除への代替アプローチがますます探求されています。
これらの強力な化学物質による土壌の酸性度も変化します
Slepetiene et al。、 2020
*必読 :土壌の種類
人為的活動は、生態系の損傷を引き起こし、土壌の健康を阻害する傾向があります。 最終的には再生不可能な資産を使い果たします。したがって、さまざまな環境にやさしい方法を採用することが不可欠です。現状では、 持続可能な農業は、私たちの現在のニーズを満たすだけでなく、健全な未来を保証する能力を提供するため、不可欠です。 従来の有害な農耕慣行では実現できない何かSantoyoet al。、 2017年
農作物生産のための農薬の使用を減らすために、 有益な根圏微生物は、持続可能な農業ソリューションに利用できます。従来の化学/合成農薬と比較して、 生物農薬にはいくつかの利点があります。 100%の生分解性と水溶性を含みます。
したがって、植物由来の生化学物質と微生物は、農業における植物病害を防除するためのより安全な代替手段です。
PGPRとは何ですか?
自由生活植物の成長促進根圏細菌(PGPR)は、植物を取り巻く根や土壌にコロニーを形成します。 彼らの成長を促進し、 発達、 と健康。
PGPRは生物的防除剤として分類することもできます。 バイオ肥料、 または生物農薬、 その活動/能力に応じて。
根圏細菌(PGPR)を促進する植物の成長は、農業における重要なプレーヤーですエテサミとマヘシュワリ、 2018年
*必読 :バイオ肥料ガイド
PGPRは、植物の健康と成長に悪影響を与える有害な有機体の拡散を制御または防止する機能を通じて、その有益な効果を発揮します。
それは窒素を固定することによって作物の幸福に役立ちます、 リン酸塩を可溶化、 重金属の削減、 植物ホルモン(オーキシンのような、 ジベレリン、 サイトカイニンなど)、 土壌有機物の鉱化作用、 作物残渣の分解、 植物病原体の抑制、 など彼ら、 2019年
病原体の生物的防除は、PGPRによって次のいくつかの方法で行われます。
1.1。 一部の細菌は、病気の原因となる病原体よりも速く、より効果的に植物のニッチにコロニーを形成することができます。これにより、有益なバクテリアが栄養素を奪い合うため、有害な生物の栄養素の利用可能性が低くなります。
2.2。 さらに、 一部の細菌は抗生物質を産生します。これらの抗体は、低濃度で病気の原因となる生物にとって致命的な有機化合物です。
3.3。 最後に、 バクテリアは植物に抵抗メカニズムを誘発します 誘導全身抵抗性(IRS) 。これにより、防御代謝物の生成がトリガーされ、植物の防御能力が向上します。

PGPRの利点
化学防除化合物よりも生物的防除剤としてPGPRを使用することには特定の利点があります。
PGPRは有益ですが、 天然に存在する微生物。それらはまた無毒で安全に使用できます。さらに、 生態学的観点から、 それらは持続可能です(長期)。
さらに、 PGPRは、さまざまなアクションモードを備えています。 抗生物質を含む、 シデロホア、 細胞壁を分解する酵素、 バイオ界面活性剤、 と揮発性物質、 植物の全身抵抗性と同様に。
非生物的および生物的植物ストレスに対するPGPR
植物にはさまざまな種類のストレスがあります。これらのストレスは、大きく2つのタイプに分類できます。 非生物的および生物的ストレス。
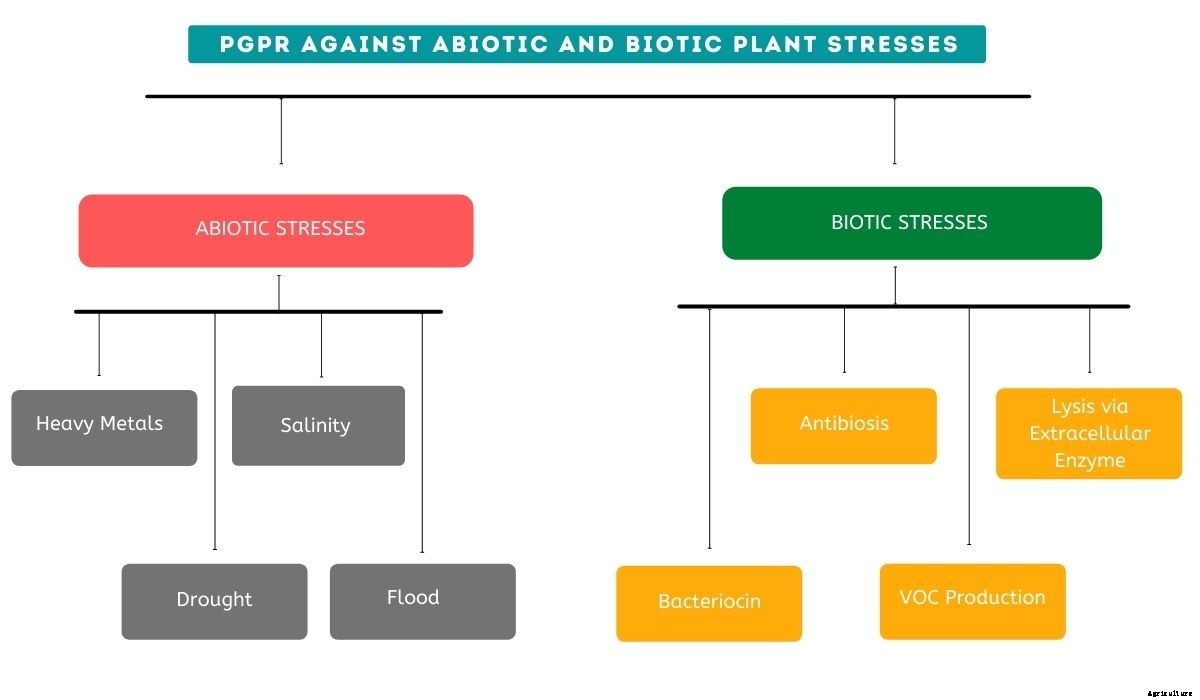
非生物的ストレス
非生物的ストレスは、微生物の多様性に影響を及ぼし、土壌の物理化学的特性も変化させる不利な環境条件によって決定される可能性があります。
非生物的ストレスの一部として、 植物の微生物叢と周囲の生態系に悪影響を与える多くの条件があります、 重金属毒性など、 塩分、 干ばつ、 と洪水。
水銀のような有害金属の場合、 アスベスト、 カドミウム、 土壌中の鉛骨材、 それらは植物のストレスを引き起こし、作物の生産性を著しく低下させます。土壌のpHとテクスチャーに悪影響を与えることに加えて、 金属の凝集は、土壌中のいくつかの生物学的プロセスに直接影響します。 作物の成長を損なう。
根圏に金属が存在すると、養分吸収が妨げられて植物の成長が遅れます。
PGPRは植物の成長と生産性を向上させますが、 また、さまざまなメカニズムを通じて土壌金属汚染物質を調整し、土壌特性を改善します。
この問題は、PGPRに金属抵抗を接種することで解決できます。誘導された誘導によるPGPRにより、植物を非生物的ストレスから効果的に保護することができます 全身耐性(IST)。
NS シデロホア 微量金属錯体を形成する微生物代謝物です。これらは、鉄との親和性が良好な低分子量化合物です。微生物は、鉄が土壌に不足しているときにそれらを生成します。金属ストレスは、微生物によって生成されるシデロホアによって効果的に抵抗されます。
バイオサーファクタント 両親媒性複合材料は主に微生物の表面に見られます。それらは微量金属の耐性を改善し、土壌金属の除去を支援します。
有機酸 のような低分子量で シュウ酸 と クエン酸 PGPRによって生成されます。これらの有機酸は、農業における金属ストレスを軽減します。 PGPRは 無機酸 それは沈殿による金属ストレスを防ぐことができます。
細胞外高分子物質(EPS) 高分子量のホモ多糖類またはヘテロ多糖類の微生物ポリマーです。根圏細菌は、リポ多糖などの細胞外多糖類を放出します。 多糖類、 溶解可能なペプチド、 糖タンパク質と陰イオン制限領域を作成し、バイオソープションによる重金属の除去または解毒を助けます。
次の表は、土壌中の重金属のPGPRバイオレメディエーションを示しています。
| PGPR | 植物 | 金属 | 栽培条件 | PGPRの役割 | 参考文献 |
Brevundimonas
ディミヌータ、
Alcaligenes faecalis | ホタルイ
ムクロナトゥス | 水星 | 温室 | •ファイトレメディエーションの増加
•土壌の毒性の低下 | ミシュラ他、 2016年 |
バチルス、
ブドウ球菌、
アエロコッカス | Prosopis juliflora、
Lolium mltiforum | クロム
カドミウム、
銅、 リード
と亜鉛 | 温室の状態 | •の効率を向上させる
ファイトレメディエーション
•高濃度を許容します。の
•クロム。 | ワニとカーン、 2012年 |
リゾビウム属、
マイコバクテリウム属 | エンドウ | クロム
(VI) | 温室の状態 | •集中力の向上
植物の窒素
•クロム毒性の低下 | ミシュラ他、 2016年 |
バチルス
メガテリウム | セイヨウアブラナ | リード | フィールド条件下で | •土壌汚染の減少
•植物の総乾物収量 | ライヒマン、 2014年 |
ブラジリゾビウム
日本住血吸虫
CB1809 | Helianthus annuus
とトリチクマ
estivum | 砒素 | ポットスタディ | •過剰な植物バイオマス
•高条件での成長
ヒ素濃度 | Yavar et al。、 2014年 |
Mesorhizobium huakuii
亜種
レンゲイB3 | トマト
ゲンゲ | カドミウム | 水耕栽培 | •PCSAt遺伝子の発現
細胞が結合する能力の増加
Cd2 | Sriprang et al。、 2003年 |
| 枯草菌SJ-101 | カラシナjuncea | ニッケル | 成長におけるポット実験
チャンバー | •の蓄積を促進しました
ニッケル。 | Zaidi et al。、 2006年 |
アゾトバクターchroococcum
HKN-5、 バチルスメガテリウム
HKP-1 B.ムシラギノサス
HKK-1 | カラシナjuncea | リード、 亜鉛 | でのポット実験
温室 | •刺激された植物の成長
•金属毒性から植物を保護しました | ウー他、 2006年 |
農業経済が塩分条件を持つことは有害です。長期の農薬使用の結果として、 塩分は時間の経過とともに土壌中に蓄積します、 これは塩分問題につながります。
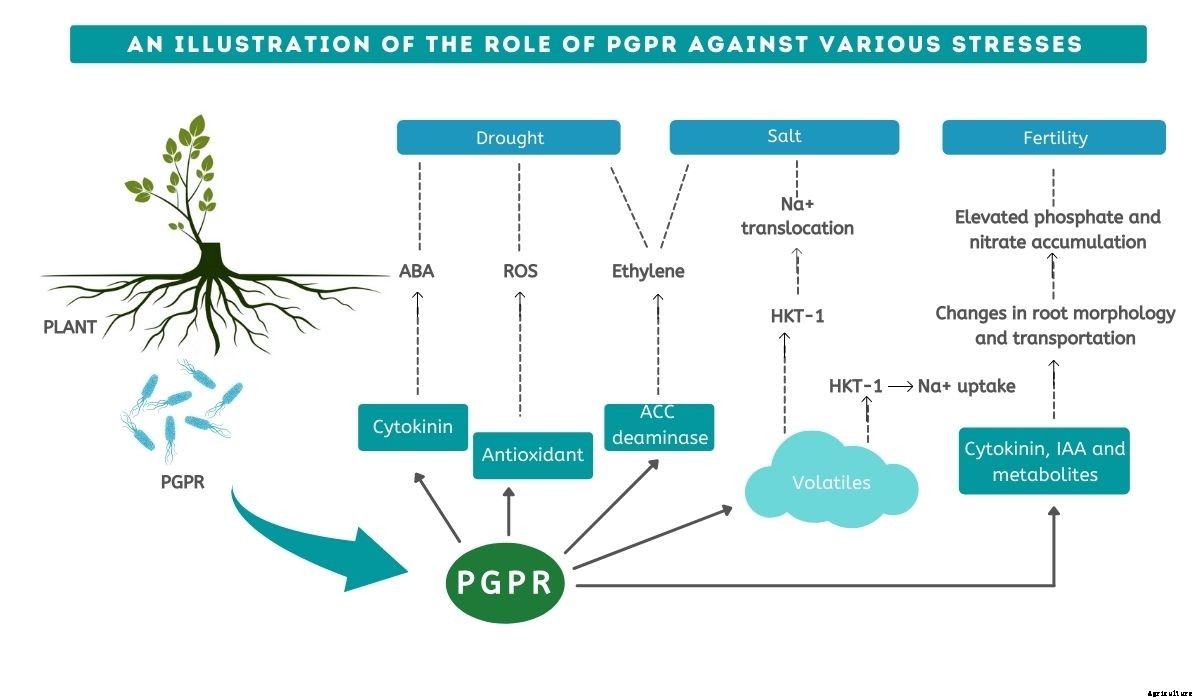
塩分ストレス下で、 活性酸素種(ROS) 含む O-2、 O2、 と H2O2 細胞を損傷し、 これは酸化ストレスとして知られています。酵素成分と非酵素成分の両方を生成できるPGPRは、植物が塩分ストレスに耐えるのに役立ちます。 H2O2レベルを管理することにより、 NS 酵素的および非酵素的抗酸化システム そのような毒性を中和します。 ROSレベルは、カタラーゼやアスコルビン酸ペルオキシダーゼなどの酵素と、アスコルビン酸などの非酵素成分によって日常的に制御されています。
PGPRプロデュース ACC(1-アミノシクロプロパン-1-カルボキシレート)デアミナーゼ、 植物を保護します エチレンストレス。
世界中の農業生産性を妨げる主な要因は干ばつです。干ばつ耐性とは、干ばつの間も持続し、耐える植物の能力を指します。
バクテリアによって引き起こされるルートアーキテクチャを変更することにより、 総根面積が増加します 、 栄養素と水分の吸収が改善され、 これは全体的な成長を促進します。バクテリアは、直径が小さく、深さが深い細根の数を増やします。 したがって、根系の総表面積が増加します。
PGPR株の接種に成功すると、 作物の生産性は ほぼ平均的なシュートの成長を維持する 干ばつストレスの間。アクセス可能な葉の表面が減少し、 水の蒸発損失が減少します 。
植物の水の状態は、 相対含水量(RWC) 。 PGPRで処理された植物は、PGPRで処理されていない植物よりもRWCの管理が優れていました。 PGPRは、乾燥した地域での植物の気孔閉鎖を調節し、植物のRWCを改善することができます。
植物では、 浸透圧の変更は、干ばつストレスに対処するための主要な適応戦略です。干ばつストレスを経験している植物では、 プロリンは重要な浸透圧調節物質です。プロリンレベルが高い植物は、干ばつに対してより耐性があります。 プロリンレベルは、PGPR接種によって増加する可能性があります。
植物の成長と発達は、化学成長調節剤と植物ホルモンのようなものによって促進されます サイトカイニン、 アブシジン酸、 ジベレリン、 オーキシン、 ジャスモン酸(JA) と エチレン 。 PGPRは、植物ホルモンと成長調節因子を調節することにより、干ばつストレスを受けた植物の発達を促進します。
洪水の間、 根系でのガス交換は減少します>これはエチレンの蓄積をもたらします。エチレンは、植物の洪水耐性形質を調節する責任があります。 ACCレベルが高いと、洪水時に根系の酸素レベルが低くなります。
根に蓄積された高濃度のACCは、ACCデアミナーゼによって減少します。 これにより、ACCが根から拡散することができます。このメカニズムにより、フラッディング中およびフラッディング後のエチレンレベルが低下します。
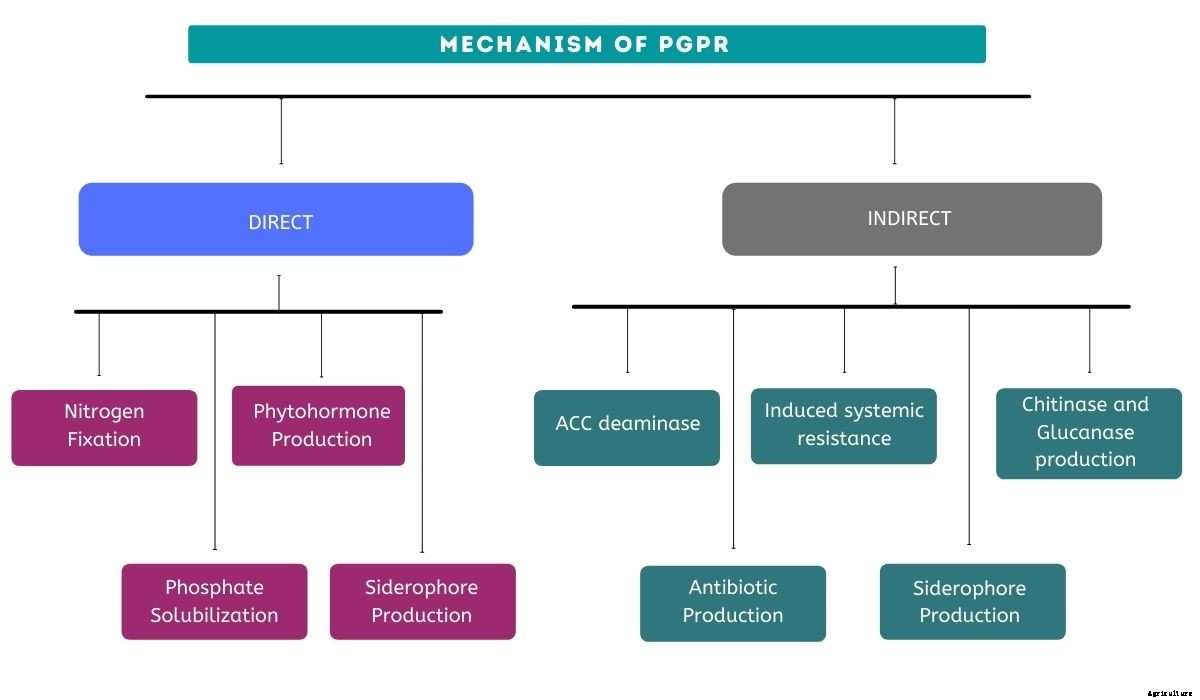
生物学的ストレス
植物では、 生物的ストレスは生物によって引き起こされます、 バクテリアなど、 ウイルス、 菌類、 昆虫、 と線虫。これらの生物は宿主の栄養素を妨害し、 植物の死をもたらします。生物的ストレスは収穫前と収穫後の両方の損失に寄与します。
病原体の生物的防除に関与する微生物はほとんどありませんが、 それでも、PGPRはバクテリオシンを含むさまざまなメカニズムに従って多くの病気からの保護を生み出すことが知られています。 抗生物質、 揮発性有機化合物(VOC)の生産、 細胞外酵素による溶解Hamidet al。、 2021年
細菌に対する細菌毒素 別名 バクテリオシン 抗菌活性が制限されたペプチド分泌物です。バクテリオシンは、グラム陰性菌の両方によって産生されます(たとえば、 コリシン )およびグラム陽性菌(例えば、 ナイシン )。実験室の条件下では、 バクテリオシンは、トマトのバクテリアスポット病と戦うのに有益であることが示されています。
それらの抗菌性のために、 殺虫剤、 抗ウィルス薬、 植物毒性、 細胞毒性、 および駆虫特性、 PGPR抗生物質は他のものより強力です。シュードモナスは、さまざまな抗真菌物質を生産しています。 2を含む 4ジアセチルフロログルシノール(2、 4-DAPG)、 ブチロラクトン、 ラムノリピッド、 N-ブチルベンゼンスルホンアミド。
特定の線虫や微生物の生物的防除の専門家であるPGPRによって分泌される多くの揮発性有機化合物(VOC)があります。 VOCにはベンゼンが含まれます。 シクロヘキサン、 テトラデカン、 および2-(ベンジルオキシ)-1-エタンアミン。 HCNは、いくつかの植物病原体を制御できるVOC(根圏微生物によって供給される)の1つです。
PGPRは、植物が感染の原因となる微生物と戦うことを可能にする溶解性化合物を生成します。根圏細菌は細胞外酵素(キチナーゼとβ-1)を生成します 細胞壁の溶解に関連する3-グルカナーゼ)。キチナーゼとβ-1 3-グルカナーゼは、壁がキチンとβ-1でできている真菌細胞上の強力な抗真菌化合物です。 4-N-アセチル-グルコサミン。
PGPRと植物の共生関係
植物と微生物の間の代謝相互作用は、アミノ酸を含むことが提案されています、 砂糖、 有機酸、 および植物によって提供される他の炭素源。
植物と根圏微生物の間の代謝関連は、このニッチで探求することができます。
根圏微生物からの生物学的代謝物は、生態学的な成功に不可欠です。それらの基質取り込みパターンによると、 この生息地の根圏微生物は重要な生態学的役割を果たしています。
多くの根圏細菌株は、天然の根系には見られない新しい代謝産物を排出するように機能します。したがって、ルートシステムを補完します。
砂糖をたっぷり加えることで、 土壌微生物は急速に増殖し、 土壌中の炭素は土壌中の限られた容量にあるという印象を与えます。したがって、 植物には、複数の代謝経路を介して外部に拡散する十分なレベルの炭素が含まれていると考えられています。
植物は主に呼吸によって炭素を固定しますが、 根圏堆積物も炭素を放出します。
根圏細菌は植物の代謝産物を生成し、 しかし、植物の根圏堆積物は、特定の細菌株を引き付け、阻害するための大きな機会を提供するさまざまな代謝物を生成します。

リゾ微生物は窒素を供給し、 リン、 植物の成長に不可欠な利用可能な形で植物に鉄を与えます。
リゾ微生物は、ACCデアミナーゼなどの植物ホルモンを産生します。 サイトカイニン、 植物の成長と発達に不可欠なインドール-3-酢酸。
PGPRの選択基準
成功するPGPR製剤の開発のために、 therhizobacterial種は、JeyarajanとNakkeeranの次の特性を備えている必要があります。 2000年
- 植物の成長を促進します
- 質量スケールでの乗算が可能である必要があります
- 高い根圏能力を備えている必要があります
- 高い競争力のある腐生能力を実証する
- 幅広い活動を実演する
- 他の根圏細菌との生態学的適合性
- 非生物的ストレス条件(熱、 放射線、 乾燥剤および酸化剤)
- 環境にやさしいはず
バイオ肥料としてのPGPR
バイオ肥料は、植物が栄養素を利用できるようにするのに役立つ有益な微生物の生きた製剤です。その生物活性を通じて、 それは土壌の健康を改善し、したがって土壌のミクロフローラを改善します。
PGPRはバイオ肥料ですか?
はい。 PGPRは バイオ肥料 。
PGPMはこのバイオ肥料の主成分です。 PGPMは、3つの主要なグループに分類できます。 つまり、 アーバスキュラー菌根菌(AMF)、 根圏細菌(PGPR)を進める植物の発達、 と窒素固定根粒菌。
PGPRはバイオ肥料として世界中で利用されています。 収量と土壌の質の向上。 PGPRがコミットされる可能性が高いため、 それは持続可能なものにつながる可能性があります アグリビジネス 。
これらのバイオ肥料は、固体および液体の形で入手できます。 液体製剤がより効果的であることがわかっています。根の接種、 種子接種、 と土壌接種は、液体製剤の3つの主要なタイプです。
Burkholderia phytofirmansバイオ肥料をライグラスの根に適用すると、 シード、 と土、 土壌接種法は、植物バイオマスの生産を改善するのに最も効率的でした。 ファイトレメディエーションと炭化水素分解Afzalet al。、 2013年
PGPRの制約
- PGPRには、その使用を制限する自然変動特性があります。 (制御された実験室条件とは対照的に)野外条件下では、 生物がどのように振る舞うかを予測することは困難です。
- 加えて、 PGPRは生きている微生物であり、 したがって、それらをフィールドで人工的に伝播する機能を持つことは、別の課題です。
- 最適化された方法での大量生産も課題です。さらに、 野外での使用も難しいまで、生物活性の耐久性と効率を維持します。
- PGPRバクテリアは土壌に非常に長く住むことができません、 そして、時間が経つにつれて、耕運機は、フィールドで彼らの人口を維持するために再接種する必要があります。
PGPRの例
PGPRの例とは何ですか?
以下は、PGPRの例と根圏におけるそれらの役割の一部です。
代表
種族 | 役割 | 関与するメカニズム | 参加している植物 | 参照 |
アグロバクテリウム
ラディオバクター | 生体保護を改善します | 抗生物質 | - | モハンラムとクマール、 2019年 |
アゾトバクター
chroococcum | アシスト
生体刺激 | ジベレリンの生産 | シリアル | 張ら、 2019年 |
| | バイオプロテクションのエイズ | シデロフォア | - | |
| Azospirillum brasilense | バイオ肥料 | リン酸塩の可溶化 | トウモロコシ(Zea mays)、 小麦(Triticum
aestivum L.)とライス(Oryza sativa) | ルーシー他、 2004年 |
| セレウス菌 | 生体保護を高める | リポペプチド | インゲンマメ(Phaseolus vulgaris) | オンゲナとジャック、 2008;
Vaikundamoorthy et al。、 2018;
橋見ほか、 2019年 |
| | | 誘発され獲得された全身
抵抗 | トマト(S. lycopersicum)
大豆(Glycine Max L.) | |
| | バイオレメディエーション | アミラーゼの生産
産業廃棄物の修復 | -
- | |
| 枯草菌 | バイオ肥料 | アンモニア合成 | トウモロコシ(Zea mays) | Ouhaibi-Ben Abdeljalil et al。、
2016; Ait-Kaki et al。、 2014;
Tahir et al。、 2017年 |
| | 生体刺激の補助 | IAAとサイトカイニンの生産を通じて | ひよこ豆(Cicer arietinum)
トマト(S. lycopersicum L.) | |
| | バイオプロテクション | リポペプチド | - | |
| | | カタラーゼ生産 | キュウリ(Cucumis sativus) | |
| | バイオレメディエーション | 生体異物の分解と
アレロケミカル | - | |
| エンテロバクターオリザエ | バイオ肥料 | 窒素固定 | マンガートとジャム(アカシアアクミネート) | Dinnage et al。、 2019年 |
フランキアcasuarinae、 NS。
非効率的、 F.イレギュラーリス、
およびF.saprophytica | 生体刺激 | サイトカイニンの生産 | - | Nouioui et al。、 2019年 |
| クレブシエラ肺炎 | バイオ肥料を助ける | 窒素固定 | トウモロコシ(Zea mays) | クアンら、 2016;シャルマ
et al。、 2019年 |
| | バイオプロテクション | 獲得および誘発された全身
抵抗 | ピーナッツ(Arachis hypogaea) | |
| メソリゾビウムロティ | バイオ肥料 | 窒素固定 | ロータス(シロイヌナズナ) | 金子ほか、 2000年 |
メチロバクテリウム
exotorquens | 生体刺激を支援します | サイトカイニン出力 | アラビドシス、 オオムギ、 トウモロコシと大豆 | 金子ほか、 2000年 |
パエニバシラス
xylanexedens | 生体保護を促進します | キチナーゼ生産 | 小麦(Triticum aestivum L.) | Verma et al。、 2016年 |
シュードモナス
緑膿菌 | バイオ肥料を支援します | リン酸塩の可溶化 | トウモロコシ(Zea mays) | 濱田ほか、 2008;
アヘマドとカーン、 2012;
Paramanandham et al。、 2017;
チェン他、 2019;ローランス
et al。、 2019年 |
| | バイオプロテクションのエイズ | アンモニア生産 | フィールドマスタード(Brassica campestris L.) | |
| | | シアン化水素の生産 | エレファントグラス(Pennisetum
プルプレウム) | |
| | アシスト
バイオレメディエーション | セルラーゼ生産 | イネ(O. sativa)、 エンドウ豆(P. sativa) | |
| | | 重金属の取り込み | アマランサス、 トマト(ナス)
lycopersicum L.) | |
リゾビウム
レグミノサラム | 生体刺激 | ジベレリン生産 | イネ(O. sativa L.) | ヤニー他、 2001年 |
| セラチア菌 | バイオプロテクション | シデロフォアの製造、 キチナーゼと
プロテアーゼ | ペポカボチャ(Poa pratensis) | Selvakumar et al。、 2008;
RathoreとGupta、 2015年 |
ブドウ球菌
スタフィロコッカス | 生体刺激 | IAAの製造 | 観賞用種 | Manzoor et al。、 2019年 |
ステノトロホモナス
リゾフィラ | バイオプロテクション | アミラーゼ合成 | トウモロコシ(Zea mays)とカノーラ(アブラナ属)
ナプス) | Ghavami et al。、 2017年 |